光合作用作為地球上生物利用太陽能的重要反應,一直是科學研究關注的重點,是植物抗逆性研究、作物高產研究的熱點。光合作用根據其反應階段可以分為基于光能吸收傳遞轉化的光反應和基于CO2同化等酶促過程的暗反應。光反應作為植物利用太陽能的原初反應,光能的吸收傳遞和轉化主要發生在植物葉片或者藻類的類囊體膜上,由光系統II(PSII)、細胞色素b6f(Cytb6f)、光系統I(PSI)、ATP合酶(ATP Synthase)共同完成,光反應會產生NADPH和ATP兩種用于暗反應的還原劑(同化力),并釋放氧氣。目前最常用的測量光合作用光反應的手段是葉綠素熒光儀法。近幾年隨著冷凍電鏡技術的發展,有關類囊體膜蛋白超級復合物結構解析的報道越來越多。在國內,中國科學院植物研究所、中國科學院生物物理研究所、清華大學的科研人員在該領域成果卓著,已經成功解析了菠菜PSII–LHCII、豌豆PSI-LHCI、玉米PSI-LHCI&LHCII、水稻PSII-PBCP、紅藻PSI-LHCR、綠藻PSII–LHCII、硅藻PSII-LHCII的晶體結構。在國際上,知名的光合作用蛋白復合物結構研究機構有:日本岡山大學理學院生物科學系,嗜熱藍藻OEC和PSII復合體結構的解析均有該單位的科學家完成;德國馬克斯·普朗生物理研究所結構生物學系,來自該單位的科學家在2018-2019年成功解析了藍藻PSI-Ferredoxin和菠菜ATP Synthase的結構;英國謝菲爾德大學的Matthew P. Johnson研究團隊則一致專注于Cytb6f的結構,先后在2014年和2019年報道了菠菜Cytb6f結構的研究結果。生理活性是光合作用的外在表現,膜蛋白結構則是生理活性的內在基礎。結構的解析可以幫助我們認識光合作用的工作原理,從而更好地利用光合作用解決人類面臨的糧食、能源、環境等問題。下面我們按照光合作用光能利用的順序簡單分析一下光合膜蛋白復合體的結構解析帶給我們的新啟示。

光捕獲是光合作用初始的重要過程。在綠色植物中,太陽能收集是由葉綠體類囊體膜中的一系列捕光復合物(色素蛋白)來實現。LHC II是葉綠體中最豐富的整合膜蛋白,以三聚體的形式存在,并與類囊體葉綠素分子的一半結合。每個LHC II單體均包含約232個氨基酸殘基的多肽,13-15個Chla和Chlb分子,3-4個類胡蘿卜素和一個緊密結合的磷脂。除了光收集功能外,LHC II在強光條件下產生的過量激發能的無輻射消散中也發揮作用。當光強度趨于飽和時,通過激發這種光保護機制,LHC II可以弱化過剩光的破壞效應。此外,LHC II還參與調節光系統II和I的激發能量分布。

2004年3月18日,Nature以封面彩圖的形式發表來自中國科學院生物物理所常文瑞課題組題為“Crystal structure of spinach major light harvesting complex at2.72 A ? resolution”的研究論文。該研究首次解析了菠菜光合作用系統Ⅱ的主要捕光復合物(LHC-Ⅱ)的晶體結構。結果發現大型R32晶胞的一個不對稱晶胞包含十個LHC-II單體。每個單體中的14個葉綠素(Chl)可以明確區分為八個Chla和六個Chlb分子。每個葉綠素的躍遷偶極矩的方向分配已經實現。所有Chlb都位于相鄰單體之間的界面周圍,并且與Chla一起是高效采光的基礎。每個單體有四個類胡蘿卜素結合位點。單體-單體界面處的葉黃素循環類胡蘿卜素可能參與了多余能量的非輻射消散,這是植物中發展起來的一種光保護策略。

LHC-II三聚體和單體中的色素布局,立體視圖顯示了LHC-II 三聚體中的色素排列模式。單體標記為I-III。為清楚起見,省略了葉綠素植物鏈和脂質。綠色,葉綠素a;藍色,葉綠素b;黃色,葉黃素;橙色,新葉黃素;品紅,葉黃素循環類胡蘿卜素。b,c,分別為基質側和腔側觀察的單體中的色素圖像。顏色名稱與a, d, e相同,分別在基質側和腔側的LHC-II三聚體內葉綠素的排列。葉綠素由三個原子表示:中心鎂原子和兩個氮原子。兩個氮原子之間的連線決定了量子躍遷偶極矩的方向。綠色,CHLA氮;藍色,CHLB氮;灰色,鎂;紫色和藍色橢圓形,近似單體面積。連接兩個葉綠素的暗線附近的洋紅色數字表示它們之間的中心到中心的距離。

基于結構的非光化學猝滅(NPQ)模型。從基質側沿二十面體C2軸的頂視圖。 DGDG顯示為黃色透明的球棒模型。葉綠素和葉黃素循環類胡蘿卜素的代表如前圖所示。黑色箭頭表示從一個三聚體到相鄰三聚體的激發能轉移途徑,橙色箭頭顯示從葉綠素Qy到葉黃素循環類胡蘿卜素S1的可能轉移途徑。紅色星號表示推定的淬滅位點。為了清楚起見,一個三聚體中的字符為黑色,而另一個三聚體中的字符為灰色。
PSII的捕光天線是一個復雜的復合物,LHC II因其在葉綠體中的豐度很高而被稱為主要的捕光復合物,位于PSII的最外圍。除此之外還有其他三個單體捕光復合物,分別稱為CP29,CP26和CP24,位于LHCII和核心復合物之間。這些是次要的捕光復合物,與LHCII相比,它們葉綠素含量相對較低。它們除了在收集太陽能并將其轉移到反應中心方面發揮作用外,這些內周天線復合物還可以在高光照條件下耗散多余的激發能。這種機制通常稱為非光化學猝滅(NPQ)。

2011年2月6日,Nature Structural & Molecular Biology雜志在線發表了中國科學院生物物理研究所常文瑞院士課題組題為“Structural insights into energy regulation of light-harvesting complex CP29 from spinach”研究論文。該研究解析了菠菜次要捕光復合物CP29的2.8分辨率晶體結構,并在晶體結構的基礎上深入分析討論了CP29的捕光、能量傳遞和光保護等功能。這是國際上首個高等植物次要捕光復合物的晶體結構。
研究發現每個CP29單體包含13個葉綠素和3個類胡蘿卜素分子,這與主要的捕光復合物LHC II有很大不同。將這13個葉綠素結合位點分配為8個葉綠素a,4個葉綠素b和1個被葉綠素a和b占據的假定混合位點。基于X射線結構分析,在CP29中構建了一個集成的色素網絡。確定為兩個特殊的色素分子簇,分別是a615-a611-a612-Lut和Vio(Zea)-a603-a609,它們可能充當潛在的能量猝滅中心以及能量轉移途徑的出入口。

CP29中的色素排列。(a)沿膜平面視圖的Chls分布。Chls由三個原子表示:中心鎂原子和兩個氮原子(NB,ND)。黃色:葉綠素a鎂;白色,葉綠素b鎂;綠色,葉綠素a氮;藍色,葉綠素 b氮。每層中相鄰的Chls,兩層中最接近的Chls和腔層中兩個簇(Chls a604-b606-b607和Chls a613-b614)中最接近的Chls用黑色虛線連接。它們的中心距離(?)用紅色數字標記。Chls分子中心的坐標是通過四個吡咯氮原子的坐標算數平均來計算的。(b–d)Lut,Vio,Neo及其附近的Chls。(e)Lut和Vio的疊加顯示出與相鄰的Chls類似的布局。綠色,在Lut附近灰色,在Vio附近。每個Chla中的氮原子NB和ND分別以藍色和品紅色表示。

強相互作用的色素團a615–a611–a612–Lut。(a,b)a611–a615的Chl對。氫鍵用暗虛線相連。Lys199與G3P之間的離子鍵用藍色虛線(a)表示。Chls a611和a615的Qy躍遷偶極矩分別用綠色和青色箭頭標記(b)。(c)色素簇a615–a611–a612–Lut的立體視圖。連接兩種色素的黑色虛線附近的數字表示色素之間的中心距離。
從LHC
II三聚體和CP29單體結構的解析我們可以得到一些啟示,CP29和LHCII的結構差異,特別是它們的色素組成和空間排列,可能是它們在光合捕光和調控過程中功能不同的原因。在CP29結構中新發現的Chl
a615與Chls
a611-a612和Lut一起,組成了一個強相互作用的色素團簇,可能是能量猝滅中心的潛在位點。在過剩光照條件下,類囊體腔中由低pH(酸化)引發的葉黃素循環(紫黃質通過花藥黃質脫環氧化成玉米黃質(Zea))被認為是NPQ中的一個重要途徑。在體內CP29中觀察到Vio向Zea的轉化效率相對較低,但在強光條件下和長時間暴露在光下時,Vio向Zea脫環氧化的程度增加。此外,低pH值下酸性殘基質子化引起的CP29構象變化對NPQ也有重要意義。
PSII的外圍天線是一個高度復雜的系統,具有很大的復雜性和多功能性。三聚體LHCII與單體CP29、CP26和CP24共同承擔著收集太陽能和光保護的重要功能。他們既相互合作,又有獨立的角色。這反映在它們在整體結構上有著明顯的相似性,卻在色素組成和排列上有著實質性的差異。CP29的三維結構為我們進一步了解PSII的小型外圍天線提供了機會。我們相信,當CP26和CP24的三維結構可用時,特別是當獲得超復雜體的整體結構(包括所有這些外圍天線)時,我們將更全面地了解PSII外圍天線的功能機制。

除了NPQ型非光化學淬滅之外,還存在一種qE型的非光化學淬滅,光系統II蛋白PsbS在qE型非光化學猝滅中起重要作用,可保護植物免受過量光照條件下的光損害。qE是由低pH值激活PsbS引發的。但是PsbS作用的機制一直不清楚。直到2015年8月10日,Nature Structural &Molecular Biology在線發表了中國科學院生物物理研究所常文瑞院士/李梅研究組題為“Crystal structures of the PsbS protein essential for photoprotection in plants”的研究論文。論文報告了菠菜中低pH下游離形式的PsbS的晶體結構,以及與qE抑制劑N,N'-二環己基碳二亞胺(DCCD)形成復合物,揭示了PsbS采用獨特的折疊模式,這不同于其他的捕光復合物超家族,它是一種非經典的色素結合蛋白。結構和生化證據表明,活性和非活性PsbS在類囊體膜中均形成同型二聚體,DCCD的結合破壞了活性PsbS二聚體的腔內分子間氫鍵。在qE期間通過低pH值激活PsbS可能涉及與PsbS二聚體的腔內分子間相互作用改變相關的構象變化相關。本研究成果從結構和生化分析為PsbS的活化和抑制機制提供了分子基礎。然而,激活的PsbS如何啟動qE尚待闡明。
已有的研究表明,在強光條件下,最初與PSII核心相關的PsbS向LHCs28遷移。如果激活的二聚體PsbS蛋白被插入到觸角之間,而沒有內部色素分子來傳遞激發能量,它可能會作為衰減器來顯著減緩LHCs之間的能量流動。此外,激活的PsbS二聚體可能直接與相鄰的LHCs相互作用,促進其構象變化,并在LHCs內部或接觸界面誘導淬滅。之前有研究人員成功的用含有PsbS、LHCII和玉米黃質的蛋白質脂質體系統體外重組淬滅過程支持了PsbS和LHC在qE期間的直接相互作用。PsbS也有可能僅僅作為pH傳感器或催化劑間接參與qE。比本研究稍早的研究表明,qE的形成與類囊體膜中PSII-LHCII超復合物的結構重組有關,并表明PsbS控制著這種重組,這可能是由低pH誘導的PsbS二聚體構象變化引起的。這里提出的PsbS結構為進一步研究PsbS在qE中的作用機理奠定了基礎。

在光合作用過程中,植物光系統II核心復合體接收來自外圍捕光復合體II(LHCII)的激發能。最初的光物理和光化學過程都是由光系統II(PSII)完成。PSII是一種嵌入類囊體膜內的超分子復合物。它包含許多蛋白質亞基和各種輔助因子,包括葉綠素、類胡蘿卜素、一個Mn4CaO5簇、一個血紅素、質體醌和脂質體。PSII的一個特征功能特性是它通過Mn4CaO5團簇催化的光誘導水氧化反應從水分子中提取電子的能力。為了收集光子能量并驅動光化學反應,植物PSII含有一系列的外圍捕光復合物(主要的捕光復合物是PSII(LHCII),次要的捕光復合物葉綠素結合蛋白(CP29、CP26和CP24))。這些天線復合物圍繞PSII的核心復合物,吸收光能并將其傳輸到反應中心,以誘導稱為P680的特殊葉綠素對中的電荷分離。然而,由于對PSII-LHCII超復合物高分辨率的結構缺乏認識,植物PSII的外周天線與核心復合物之間激發能量傳遞的精確途徑仍不清楚。此外,植物PSII中三個重要的外源亞基(PsbO、PsbP和PsbQ)的功能和相互作用尚不清楚。為此中科院生物物理所柳振峰團隊通過單顆粒冷凍電鏡技術,在3.2 ?分辨率下解析了1.1兆道爾頓的菠菜光系統II–LHCII超級復合物的結構。首次揭示了LHCII、CP29以及CP26向核心天線復合物CP43或CP47傳遞能量的途徑,對于進一步在分子水平理解PSII-LHCII超級復合物中的能量傳遞時間動力學和光保護機理具有重要意義。相關結果于2016年5月18日發表在Nature雜志上,題目為 “Structure of spinach photosystem II–LHCII supercomplex at 3.2? resolution”,電鏡結果顯示PSII-LHCII是一個同源二聚體的超分子體系,其中每個單體含有25個蛋白亞基,105個葉綠素分子,28個類胡蘿卜素分子和其他輔因子。三個外部亞基(PsbO, PsbP 和 PsbQ)對于光系統II的最佳放氧活性至關重要,它們形成了一個三角形冠狀結構,環繞著CP43和D1蛋白的Mn4CaO5結合結構域(光系統II中的1個Ca原子與4個 Mn原子由5個O原子作為橋連成Mn4CaO5簇,是光催化水分子斷裂生成氧氣的放氧復合體的核心)。一個主要的三聚體LHCII和兩個次要的單體LHCII分別與核心復合物的每一個單體結合,且三個小的內部亞基(PsbW, PsbH 和 PsbZ)加強了天線與核心復合物間的相互作用。通過分析緊密連接的界面間葉綠素分子,我們對天線與核心復合物間的能量傳遞通路有了更深入的了解。

菠菜PSII-LHCII超復合物的總體結構。a,b,菠菜C2S2型PSII-LHCII超復合物的結構。a,沿膜從基質側水平角度觀察。b,沿膜垂直角度的側視圖。虛線表示兩種單體之間的估計界面區域。主要組件顯示為不同顏色的卡通和棒狀模型,而12個小的內在亞基顯示為黃色球形模型。c,卡通圖,顯示了超復合物的膜嵌入亞基的分配。僅顯示一種單體,并且顏色代碼與a中一致。d,超復合體的腔側暴露區域。該視圖是沿腔從腔側面觀察的。
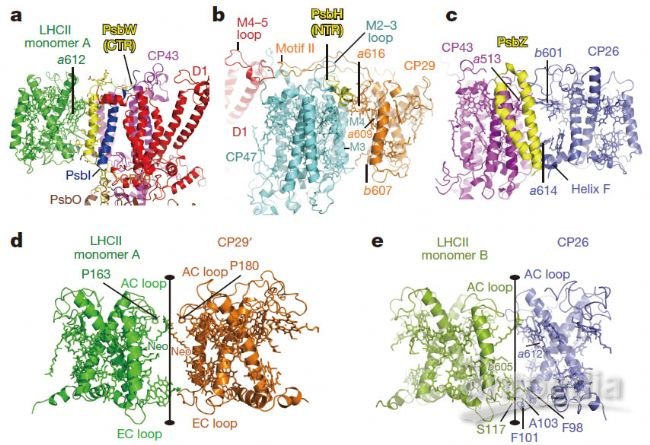
PSII-LHCII超復合物中的天線-核心和天線-天線相互作用。a,LHCII和CP43 / D1之間的接口。它們的相互作用由PsbW和PsbI介導。 PsbW(CTR),PsbW亞基的C端區域。b,CP29和CP47 / D1之間的相互作用。 M4–5環,D1的第四個和第五個跨膜螺旋之間的環區域;M3和M4,分別是CP47的第三和第四跨膜螺旋;M2–3環,CP47的第二個和第三個跨膜螺旋之間的環;PsbH(NTR),PsbH亞基的N端區域。c,CP26和CP43 / PsbZ之間的相互作用。d,LHCII和CP29'之間的接觸。e,CP26–LHCII相互作用。

從天線復合體到反應中心的能量轉移途徑。a,PSII-LHCII超復合物中葉綠素的分布模式。Chla和Chl b分別用綠色和藍色區分。箭頭指示LHCII和CP29'/ CP26(紅色),LHCII / CP29'/ CP26和核心天線(CP43 / CP47')(洋紅色),CP43 / CP47'和D1 / D2'反應中心之間的潛在能量轉移路徑(青色)。五個紅色橢圓形表示在CP29'中Chla611-Chla612對周圍的潛在能量猝滅位點,黑色橢圓形表示在CP29'中Chla603-Chla609對周圍的另一個潛在淬滅位點。這些淬滅位點可能在強光條件下被激活,以消散有害的過剩能量。虛線表示每個單獨復合物的近似邊界。 b–d,界面葉綠素支持LHCII和CP29'(b),LHCII和CP26(c)以及LHCII和CP43(d)之間的能量轉移。 e,f,CP29和CP47(e)之間以及CP26和CP43(f)之間的界面處的葉綠素。虛線附近的數字表示兩個相鄰葉綠素之間的Mg至Mg距離(?)。界面葉綠素以洋紅色突出顯示。
綜上所述,我們可以深入了解PSII中的能量傳遞途徑。通過位于周圍和核心天線復合體之間的界面葉綠素對,激發能可以從LHCII,CP26和CP29傳遞到CP43或CP47。LHCII三聚體在其單體-單體界面處包含三個富含Chb的簇,其中兩個簇連接到CP26和CP29'。由于Chlb的能量水平高于Chla,LHCII和CP26之間(或LHCII和CP29'之間)的能量轉移可能會從LHCII的富含Chlb的區域流到CP26(或CP29')中與之相鄰的富含Chla的區域。對于LHCII和CP29'之間的能量傳遞,來自腔側面的Chl b605monomerA/LHCII和Chl a604 / b606CP29'形成最接近的內復合物對,Mg-Mg距離(Dcentre)為17.7 / 18.6?,而在基質層Chlb608monomerA/LHCII和Chlb608CP29'連接的Dcentre為23.0?。因此,CP29'的富含Chlb的區域與LHCII的區域相連,促進了二聚體超復合物的兩個相鄰單體之間的能量轉移,大概是通過Chlb605monomerA/LHCII 到 Chla604 / b606CP29'和b608monomerA / LHCII到b608CP29'的路徑。對于LHCII和CP26之間的能量傳遞,來自LHCII三聚體的單體B的Chlb608連接到CP26的Chla612和Chla610,連接Dcentre分別為21.8和21.9?,而來自腔側面的LHCII的Chlb605可能轉移其激發能量到CP26的Chla604中,因為它們的連接Dcentre為19.3?。
LHCII中最低能態的葉綠素為Chl
a610,a612和a611簇,被稱為末端發射域。LHCII三聚體中平衡的激發能將集中在該簇上。在超復合物中,來自LHCII三聚體的單體A的末端發射體可能會將其能量傳輸到Chla506CP43,該基團相對于Chla611monomerA/LHCII位于有利的方向(幾乎平行)和距離(17.1?
Dcentre)。在Chla611下方,來自腔側面的LHCII的Chla614以25.1?的Dcentre連接到Chla501CP43。這兩個途徑構成了LHCII和核心復合物之間能量轉移的基礎。在沒有次要天線復合體的情況下,LHCII可以將能量直接轉移至核心復合體,但是在次要天線敲除的突變體植物中,LHCII和PSII核心之間的功能連接受到嚴重損害。
為了在CP29和CP47之間進行能量轉移,將Ch1a616CP29夾在Ch1a609CP29和Ch1a616CP47之間,Dcentre分別為9.3 ?和14.4 ?。Ch1a616CP29和Ch1a609CP29 / a616CP47之間最接近的邊對邊距離(Dedge)為3.4 / 4.2?,表明這些葉綠素形成了強耦合對。Ch1a616CP29在間隙中的位置使其成為關鍵的連接子,將激發能從CP29傳遞到CP47。此外,CP29的Chla603是以18.6? 的Dcentre直接連接到Chla610CP47的。在腔側面,能量可能會以19.1 ? 的Dcentre從Ch1b607CP29轉移到Ch1a607CP47。或者Ch1b607CP29將它的能量傳遞給Ch1a603-a609CP29,然后能量進一步由Chl a616CP29 釋放給 Chl a616CP47。在這些潛在途徑中,Ch1a616CP29至Ch1a616CP47可能是CP29和CP47之間最有效的能量轉移途徑,因為這兩個葉綠素在界面上配對最緊密。
CP26與CP43相互作用密切,它們之間的能量轉移可能通過多種潛在途徑發生。Ch1a611CP26與最紅的Ch1a612形成一個強耦合對,該耦合對可能是CP26的終端發射域。Chla611以18.2? 的Dcentre連接到Ch1a512CP43,而以17.2? 的Dcentre連接到Chla513CP43。同時,Chlb601CP26以4.8 ? 的Dedge(Dcentre為12.6 ?)與Chla513CP43耦合,并且還以19.2 ? 的Dcentre連接到Chla512CP43。在腔側面,來自Ch1a614CP26的激發能量可能主要被16.0 ?Dcentre 處的Ch1a503CP43吸收。因此,從CP26傳輸的能量將在基質側被Chla513–Chla512接收,或者被CP43腔側面的Chla503接收。
當外圍天線復合體的激發能被核心天線復合體收集,隨后的能量通過位于CP43,CP47,D1和D2內的Chla網絡從CP47或CP43傳遞到P680特殊葉綠素分子對。在強光條件下,主要和次要LHCII中的色素分子簇可充當非光化學猝滅位點,將有害的多余激發能耗散為熱量。超復合體內的潛在猝滅位點主要位于相鄰天線復合體之間的界面處或附近。這些位置非常適合它們在到達反應中心之前攔截和消耗多余的能量。比該研究稍早一些時候的報道顯示,生物物理模型研究已經獲得了有關PSII-LHCII超復合物中光收集動力學的初步信息。現在,菠菜PSII-LHCII超復合物的冷凍電鏡結構為其高度復雜的色素網絡提供了詳細的框架,并使人們能夠更深入地了解超復合物中光捕獲過程的動力學和調控。
由于相關研究內容非常專業,難免有些理解不準確或者編輯整理的疏漏,請以英文原文為準。
參考文獻
1. Liu Z, Yan H, Wang K, et al. Crystal structureof spinach major light-harvesting complex at 2.72 ? resolution. Nature,2004, 428(6980): 287.
2. Pan X, Li M, Wan T, et al. Structural insights into energy regulation of light-harvesting complex CP29 from spinach. Nature structural & molecular biology, 2011, 18(3): 309.
3. Fan M, Li M, Liu Z, et al. Crystal structures of the PsbS protein essential for photoprotection in plants. Nature structural & molecular biology, 2015, 22(9): 729.
4. Wei X, Su X, Cao P, et al. Structure of spinach photosystem II–LHCII supercomplex at 3.2 ? resolution. Nature, 2016, 534(7605): 69.